| 21世纪最有生命力的行业之一,不得不说有生物行业。生物行业的发展速度已经超过了以往任何的时代,而且越来越加深入的研究着生命这个永恒不变的主题。目前随着对于表观遗传学的研究逐步深入,越来越多新的发现被生物界人士所震掠。其中新“四大碱基”的发现,对于生物行业的研究者来说,是又惊又喜。惊的是我们对于生命的认识,还有很长的路要走;喜的是我们对于生命的认识又向前迈进了一大步。相信表观遗传学研究的更加深入会给生物科学的发展,注入更多的活力;坚信越来越多的研究人员,会关注并研究这一课题。
下面我司经过整理相关的文献,简要介绍“新四大碱基”:
新四大碱基是什么?
几十年来,科学家们一直认为DNA中只包含有4种碱基:鸟嘌呤(G)、胸腺嘧啶(T)、腺嘌呤(A)、和胞嘧啶(C),这4种碱基已成为我们对基因代码如何形成生命的认识的基础。此处对于“新四大碱基”的说法主要是针对传统研究的四种碱基而言的。具体如下:
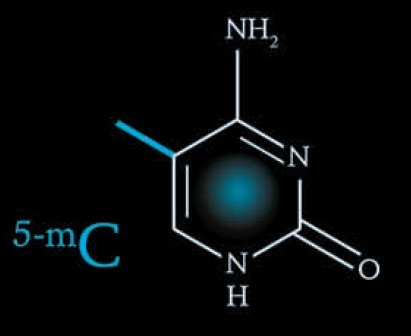
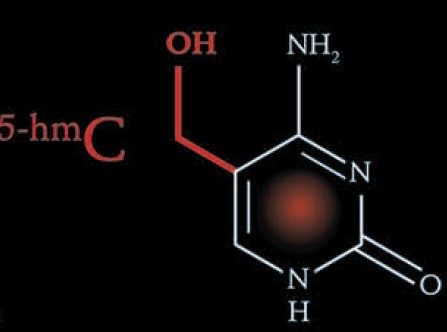
第5种碱基:5-甲基胞嘧啶 (5-methylcytosine) 第6种碱基:5-羟甲基胞嘧啶 (5-hydroxymethylcytosine)
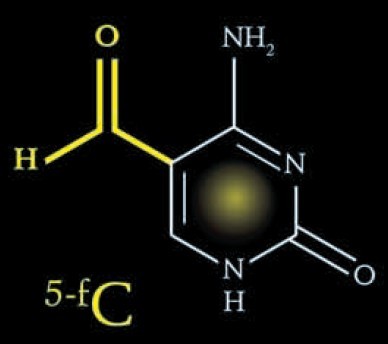
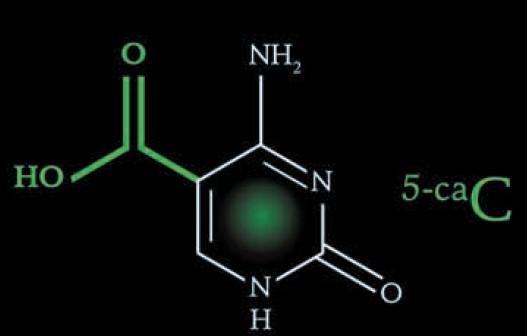
第7种碱基:5-甲酰基胞嘧啶 (5-formylcytosine) 第8种碱基:5-羧基胞嘧啶 (5-carboxlcytosine)
对于研究者的几点启示:
第5种碱基的意义:
5-甲基胞嘧啶(5-methylcytosine, 5mC)被认为是哺乳动物基因组中除腺嘌呤(adenine)、胸腺嘧啶(thymine)、胞嘧啶(cytosine)及鸟嘌呤(guanine)之外的第五种碱基。它与染色质的另外一种重要组分组蛋白及其翻译后修饰的组合决定了特定基因组区域染色质的结构及基因转录活性,从而形成了一层叠加于碱基序列上的表观遗传信息。胞嘧啶甲基化,也称为DNA甲基化,参与了诸多生物学过程,包括基因印迹、X染色体的失活、基因组稳定性的维持、转座子及逆转录转座子的沉默及组织特异性基因的沉默等[1-2]。
在哺乳动物的个体发育中,DNA甲基化谱式主要经历了两次大规模的重编程过程,一次发生在从受精至着床的早期胚胎发育时期,另一次发生在配子发生过程中[3-4]。这两次重编程都涉及了基因组范围的主动去甲基化反应(global demethylation)。相对于基因组范围内的大规模主动去甲基化,在体细胞中会发生局部的、高度位点特异性的主动去甲基化[5-6]。DNA的去甲基化与DNA甲基化这两个过程相互平衡,维持了DNA甲基化谱式的稳定。任何一方的失调都会导致DNA甲基化谱式的紊乱,进而引起多种神经退行性疾病、免疫系统疾病以及癌症[7-8]。特别是两次基因组范围的主动去甲基化事件对于生命的起始以及生命的传承具有非常重要的意义。
DNA甲基化是由DNA甲基转移酶(DNA methyl-transferase)催化完成,那DNA去甲基化是如何完成的呢?也存在类似于DNA甲基转移酶的DNA去甲基化酶(DNA demethylase),抑或是其他的机制呢?就此,著名的华人生物学家、美国科学院院士朱健康教授在一篇综述中提出以下几种DNA去甲基化的可能方式 (图1) [5]。(1)存在特异性识别并切除5mC的糖苷酶(glycosylase),切除5mC产生AP位点(apurinic/apyrimidinic site),进而启动碱基切除修复途径(base excision repair, BER),用没有修饰的胞嘧啶取代原有的甲基胞嘧啶,最终完成DNA的去甲基化。(2)存在特异性识别5mC的脱氨酶(deaminase),通过脱氨作用将5mC转变成胸腺嘧啶,形成G/T错配。进而由识别G/T错配的糖苷酶,如TDG、MBD4等,启动BER途径,完成DNA的去甲基化。(3)通过核苷酸切除修复途径(nucleotide excision repair, NER)切除含有5mC的一段DNA,并用含有未修饰胞嘧啶的同样序列进行替换,进而完成DNA的去甲基化。(4)通过氧化作用将甲基基团氧化成羧基基团,再通过脱羧作用完成DNA去甲基化。(5)通过水解作用直接将甲基基团去掉。朱健康教授实验室发现在拟南芥中存在着一种由DNA 糖苷酶(DME、DML2、DML3和ROS1等)介导的主动去甲基化机制。ROS1等可以切除5-甲基胞嘧啶,启动碱基切除-修复途径(base excision repair, BER)完成DNA去甲基化[5]。但在哺乳动物中并没有鉴定到这些糖苷酶的同源蛋白,提示哺乳动物中可能存在不同的DNA去甲基化机制。
长期以来,DNA主动去甲基化的机制一直是表观遗传领域的一个最基本的问题。但由于DNA主动去甲基化事件只限于卵子受精后分裂前和生殖细胞生成过程,时间窗口很短,细胞来源有限,无法获得足够数量的细胞或组织进行分子生物学操作来鉴定基因,更无法进行生化操作去纯化酶,因此研究起来极其困难。但这个难题从来没有缺少过关注,很多实验室试图用其他方法来解决这个问题,也包括本实验室。1999年,加拿大McGill大学的犹太裔生物学家Moshe Szyf教授在Nature上发表文章,声称他们实验室鉴定到了DNA去甲基化酶,这个酶含有甲基化CpG结合结构域,能够在体外将5mC转变成C[9]。这在表观遗传领域引起了一时的轰动,但其他实验室重复不出来这个结果。直到2007年,德国海德堡的Christof Niehrs实验室利用大规模筛选的方法鉴定能够重新激活被DNA甲基化沉默的荧光素酶(luciferase)报告基因的蛋白。他们发现DNA损伤诱导蛋白Gadd45a(growth arrested and DNA-damage-inducible protein 45 alpha)能够促进修复途径介导的DNA去甲基化[10]。但Gadd45a基因敲除并不妨碍胚胎发育,也不影响受精卵分裂前雄原核的主动去甲基化过程。另外,Gadd45a只是启动一种DNA修复机制,并不是真正的去甲基化酶。2008年,美国犹他大学的David Jones实验室与Bradley Cairms实验室合作,将甲基化的DNA注射入斑马鱼的胚胎中来研究DNA去甲基化。他们提出脱氨酶AID (activation induced deaminase)、糖苷酶MBD4及Gadd45a蛋白相互协作,通过脱氨偶联的碱基切除修复途径完成DNA去甲基化[11]。2010年,英国剑桥大学的Wolf Reik教授实验室报道AID蛋白的缺失影响了原始生殖细胞中的去甲基化事件[12]。这为AID蛋白通过脱氨作用参与DNA去甲基化提供了进一步的证据。但还是不能完全解释基因组范围的主动去甲基化事件。在2009年,DNA去甲基化研究出现了一个全新的突破。美国科学院院士Anjana Rao实验室发现DNA双加氧酶Tet1蛋白在体外可以将5-甲基胞嘧啶氧化成5-羟甲基胞嘧啶(5-hydromethylcytosine, 5hmC)[13],且5hmC稳定存在于胚胎干细胞及浦肯野细胞中[14],这提示5mC可能通过氧化途径经由5hmC最终转化成非甲基化的胞嘧啶[6]。
在粗糙脉胞菌(Neurospora crassa)中存在一种尿嘧啶代谢补偿途径。胸腺嘧啶在胸腺嘧啶水解酶(thymine-7-hydrolyase,T7H)的催化下经三步氧化反应被转变成5-羧基尿嘧啶,随后5-羧基胞嘧啶在异乳清酸脱羧酶(iso-orotate decarboxylase, IDCase)的作用下转变成尿嘧啶[6,15]。那Tet蛋白是否具有类似于T7H的功能,通过多步氧化反应将5mC转变成5-羧基胞嘧啶(5-carboxylcytosine, 5caC)呢?本实验室正是在这个假说的推动下开始了哺乳动物DNA主动去甲基化机制的研究。首先,优化了Tet蛋白的体外反应条件,除了加入辅因子2-OG (2-oxoglutarate)及二价铁离子,还加入了1 mmol/L ATP。高效液相色谱(high performance liquid chroma-tography, HPLC)及薄层层析(thin layer chromatography, TLC)的结果显示5mC被转变成一种新的形式。经质谱鉴定,这种新的修饰形式为5caC。在293T细胞中过表达Flag-Tet2蛋白可在基因组DNA中检测到5caC,而Tet2突变体蛋白则不能产生5caC。这些结果表明Tet蛋白确实可以通过氧化作用将5mC转变成5caC。接下去的问题是5caC如何转变成非甲基化的胞嘧啶的呢?是存在类似于IDCase的5caC脱羧酶,还是其他途径,比如碱基切除修复途径呢?2010年,英国剑桥大学的Azim Surani实验室的研究工作显示碱基切除修复途径参与了小鼠生殖系的重编程[16]。糖苷酶是BER途径最上游的蛋白分子。本实验室纯化了目前已知的四种针对G/U错配的糖苷酶,即TDG、MBD4、UNG1及SMUG1,并检测这些糖苷酶是否可以特异性地切除5caC,结果显示这四种糖苷酶中只有TDG可以特异性地切除5caC。用anti-TDG抗体免疫去除ES核抽提物中内源的TDG蛋白导致核抽提物中5caC切除活性的大大降低。而共转染TDG和Tet2则可以消除基因组中Tet催化产生的5caC,丧失酶活的TDG突变体蛋白则没有这种作用。Tdg knockdown的ES细胞或者Tdg knockout的iPS细胞的核抽提物中5caC切除活性基本消失,同时在这两种细胞中均可以用HPLC-MS-QQQ的方法检测到5caC的积累。上述结果证明5mC可在双加氧酶Tet的催化下转变成5caC,而5caC则进一步通过TDG介导的BER途径转变成未甲基化的胞嘧啶[17]。在本实验室开展Tet蛋白通过氧化作用将5hmC转变成5caC工作的同时,美国北卡罗来纳大学、霍华德休斯医学研究所的著名华人科学家张毅教授实验室也发现了同样的生化机制。他们发现Tet蛋白可以在体外将5hmC顺序氧化成5-醛基胞嘧啶(5-formylcytosine, 5fC)和5caC,并用质谱方法在ES细胞及多种小鼠组织中检测到5fC和5caC的存在[18]。但他们没有继续深入研究5caC如何转变成未修饰的胞嘧啶。所以,本实验室首次相对完整地提出了哺乳动物DNA主动去甲基化的一种全新分子机制(图2)。
第8种碱基的意义:
1.中科院上海生命科学研究院生物化学与细胞生物学研究所徐国良研究组近日在《科学》杂志发表论文,初步阐明了DNA去甲基化的机制与过程,宣告人类发现了这个生命编码过程中的“新单词”。这意味着人类对自身受精卵变为胚胎细胞的内在机制有了更深理解。
甲基化是核酸的一种重要修饰,调节基因的表达和关闭,与动植物生长发育、疾病发生发展有密切关系。以往研究已知,DNA四大碱基还有两种修饰形式,即第5种碱基5mC、第6种碱基5hmC,它们均包含甲基(m)这个“词缀”,是甲基化的碱基。DNA甲基化会关闭某些基因的活性;而反之,DNA去甲基化则会诱导基因的重新活化及功能表达。在生殖细胞形成、受精卵向胚胎发育过程中,就出现大规模的全基因组去甲基化,成为非常重要的“生命编码”环节。其中艾德科技提供的表观遗传学产品线目前可以解决研究者对于DNA甲基化与DNA羟甲基化的研究需要。
徐国良研究组及其合作方通过研究发现,无论是体外的细胞还是培养的细胞,DNA第5、第6种碱基可以进一步形成第8种碱基5caC。这种新的修饰碱基会被自动识别出来,并从基因组中切除,再通过DNA的剪切修复,替换为没有修饰成分的胞嘧啶。
专家认为,此项研究的重要意义在于,发现了第8种碱基作为中间状态确实存在,还锁定了参与其中的特定的酶,这为DNA去甲基化机制研究提供了生物化学证据,也为下一步研究指明了方向。
研究表明,若掌握基因的使用规律,将对人类认识动植物生长发育、疾病的发生发展以及将已经分化成熟的体细胞“返老还童”为干细胞,都具有十分重要的意义。
在哺乳动物的表观遗传调控中,TET蛋白参与了DNA的主动去甲基化过程,该蛋白催化产生的5-羧基胞嘧啶可能通过两种途径被转变为胞嘧啶,其中一种途径就是通过潜在的DNA脱羧酶直接催化5-羧基胞嘧啶发生脱羧反应。由于胞嘧啶和尿嘧啶的化学结构和性质比较相似,因此,潜在的DNA脱羧酶可能与真菌中尿嘧啶脱羧酶IDCase在结构和催化机制上存在一定的相似性。
2. 2013年8月6日国际知名杂志《细胞研究》 (Cell Research)在线发表了生物化学与细胞生物学研究所丁建平组关于尿嘧啶脱羧酶的催化机制的最新研究成果,揭示了IDCase的催化机制,为寻找哺乳动物中潜在的DNA脱羧酶提供了新思路。
DNA胞嘧啶的甲基化是一种重要的表观遗传修饰,在很多生物学过程中都发挥重要作用。DNA胞嘧啶的甲基化修饰由DNA甲基转移酶催化发生,然而DNA的主动去甲基化是如何发生的却长期未有定论。近期研究表明,TET蛋白通过逐步氧化5-甲基胞嘧啶生成5-羧基胞嘧啶,且可能存在DNA脱羧酶直接催化5-羧基胞嘧啶脱羧生成无修饰的胞嘧啶。胞嘧啶和尿嘧啶的结构非常相似,此外哺乳动物中TET蛋白催化5-甲基胞嘧啶发生三步氧化反应最终生成5-羧基胞嘧啶的过程与真菌中T7H催化胸腺嘧啶转变为5-羧基尿嘧啶的过程极为相似。鉴于此,哺乳动物中很有可能存在DNA脱羧酶,而这种潜在的DNA脱羧酶与真菌中催化5-羧基尿嘧啶转变为尿嘧啶的IDCase在序列、整体结构、底物结合和催化机制等方面可能存在较多相似性。此外,IDCase的底物识别和催化反应机制也未有报道。
丁建平研究组的博士生徐曙彤和李文婧等人解析了Cordyceps militaris来源的尿嘧啶脱羧酶IDCase (CmIDCase)的野生型和突变体原酶(apo)形式以及与底物5-羧基尿嘧啶、底物类似物5-硝基尿嘧啶和产物尿嘧啶的复合物的晶体结构,以及Metarhizium anisopliae来源的IDCase的原酶形式的晶体结构。结构分析表明,IDCase呈现典型的氨基水解酶超家族所含有的(β/α)8桶状折叠结构,在原酶形式和结合配体形式的CmIDCase结构中,活性位点处都结合了一个Zn2+。进一步的突变体和酶活实验验证了参与金属离子结合和底物结合的关键氨基酸在催化反应中的功能。基于结构分析和体外生化实验结果,提出了一种新的脱羧反应催化机制。他们的研究结果不仅揭示了尿嘧啶脱羧酶IDCases的底物识别和催化机制,并为寻找哺乳动物中潜在的DNA脱羧酶提供了新思路和重要信息。
待上传中...
[参 考 文 献]
[1] Jaenisch R, Bird A. Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals. Nat Genet, 2003, 33 (Suppl): 245-54
[2] Goll MG, Bestor TH. Eukaryotic cytosine methyl-transferases. Annu Rev Biochem, 2005, 74: 481-514
[3] Reik W, Dean W, Walter J. Epigenetic reprogramming in mammalian development. Science, 2001, 293: 1089-93
[4] Reik W. Stability and flexibility of epigenetic gene regulation in mammalian development. Nature, 2007, 447: 425-32
[5] Zhu JK. Active DNA demethylation mediated by DNA glycosylases. Annu Rev Genet, 2009, 43: 143-66
[6] Wu SC, Zhang Y. Active DNA demethylation: many roads lead to Rome. Nat Rev Mol Cell Biol, 2010, 11: 607-20
[7] Robertson KD, Wolffe AP. DNA methylation in health and disease. Nat Rev Genet, 2000, 1: 11-9
[8] Robertson KD. DNA methylation and human disease. Nat Rev Genet, 2005, 6: 597-610
[9] Bhattacharya SK, Ramchandani S, Cervoni N, et al. A mammalian protein with specific demethylase activity for mCpG DNA. Nature, 1999, 397: 579-83
[10] Barreto G, Schäfer A, Marhold J, et al. Gadd45a promotes epigenetic gene activation by repair-mediated DNA demethylation. Nature, 2007, 445: 671-5
[11] Rai K, Huggins IJ, James SR, et al. DNA demethylation in zebrafish involves the coupling of a deaminase, a glycosylase, and gadd45. Cell, 2008, 135: 1201-12
[12] Popp C, Dean W, Feng S, et al. Genome-wide erasure of DNA methylation in mouse primordial germ cells is affected by AID deficiency. Nature, 2010, 463: 1101-5
[13] Tahiliani M, Koh KP, Shen Y, et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mamma-lian DNA by MLL partner TET1. Science, 2009, 324: 930-5
[14] Kriaucionis S, Heintz N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science, 2009, 324: 929-30
[15] Smiley JA, Kundracik M, Landfried DA, et al. Genes of the thymidine salvage pathway: thymine-7-hydroxylase from a Rhodotorula glutinis cDNA library and iso-orotate decarboxylase from Neurospora crassa. Biochim Biophys Acta, 2005, 1723: 256-64
[16] Hajkova P, Jeffries SJ, Lee C, et al. Genome-wide reprogram-ming in the mouse germ line entails the base excision repair pathway. Science, 2010, 329: 78-82
[17] He YF, Li BZ, Li Z, Liu P, et al. Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA. Science, 2011, 333: 1303-7
[18] Ito S, Shen L, Dai Q, et al. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxy-lcytosine. Science, 2011, 333: 1300-3
定购和索要更多qMSP、MSP、BSP、MS-MLPA和RT-PCR相关产品与技术资料,请您加QQ:1513545070
作者:Admin 上传日期:2012年04月29日 |